This transatlantic meta-analysis showed that clear-cut harvest in boreal forests consistently resulted in divergence of community composition away from that of unharvested reference forests. There was evidence of resilience for most biotic groups and forest types. Modelled times to ‘full recovery’ were ~12 to 20 years in a few cases but more often ~25–35 years, or longer than the time period for which we had data. Indeed, in several cases predicted ‘full recovery’ took >55 and up to ~85, ~95 or >100 years.
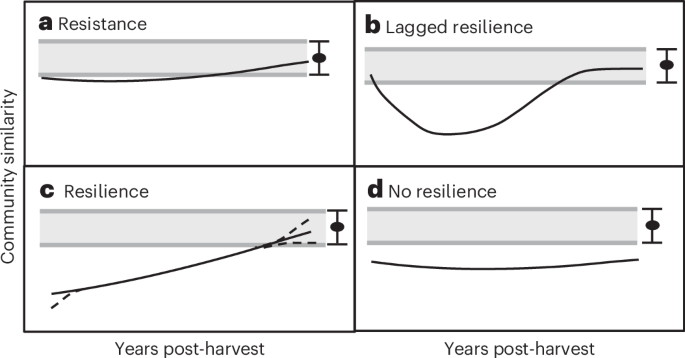
As hypothesized, post-harvest recovery of community composition varied among biotic groups and forest types and was usually nonlinear, often showing a temporal lag (initial gradual decline in harvested versus reference community similarity with a subsequent increase). Our results support previous evidence for the strong effects of clear-cut forest harvest on biodiversity6,21,24,27,28. Substantial changes in the complement of species and their relative abundances after clear-cutting, at least in the short- or mid-term, were illustrated by the dramatic post-harvest divergence from the reference community composition. Interestingly, several of our modelled times to ‘full recovery’ correspond with the ~30–40 years reported for a variety of ecosystem function and biotic variables in forests globally7, although that estimate was based on the authors’ judgement on whether a system had recovered. Similarly for lowland tropical forests redeveloping after agricultural use, a recent study suggested that community composition for several biotic groups attained 75% similarity to old-growth forests within 30 years (ref. 14).
Variation in recovery patterns
Natural disturbances create forest landscapes with a mosaic of stand ages and types, each of which is important for biodiversity12,31. Many studies have demonstrated substantive differences in post-disturbance structure, function, and biodiversity between harvested forests and those following natural disturbances such as fire26,31. We chose to compare harvested forests to the mature forest reference condition because we consider harvesting to be an additional disturbance footprint.
The recovery patterns and predicted times to ‘full recovery’ we present are generalized expected responses, with much unexplained variation around the GAMM curves. Studies exploring the influence of modifying factors and contextual differences on responses would be most welcome (for example, forest site types, natural-disturbance regimes, post-harvest forest development, condition of surrounding landscape). The GAMM curve was particularly sensitive to cases with few data for intermediate time periods (for example, beetles in mixed and broadleaf forests) and estimates of ‘full recovery’ are necessarily imprecise because we had relatively few data for >30 years post harvest (Supplementary Table 4). Studies with data for multiple years post harvest were most often chronosequences. More datasets with long-term repeated measurements for >30 years post harvest would help refine estimates of ‘full recovery’ and further improve our understanding of longer-term effects of clear-cut harvest21.
In the vast majority of cases there was some evidence of resilience (resilience or lagged resilience) with a pattern of increasing community similarity between harvest and reference over time post harvest. This is in concordance with evidence of recovery for productivity, nutrient cycling and tree and understory plant diversity following harvest26 and may be particular to the boreal forest, which often experiences a natural-disturbance regime dominated by stand-initiating disturbances such as fire. There was considerable variation, however, in the timing of recovery and the underlying data included many individual examples that showed no evidence of recovery even 50–100 years post harvest (that is, harvest vs reference similarity values ~ 0; Supplementary Fig. 1b,h–j: all beetles, bryophytes, lichens and vascular plants in conifer forests). Very poor resilience for individual sites can probably be explained by composition and structure of the post-harvest forest, which strongly reflects harvesting practices and silvicultural treatments12,18,32. Recovery was probably facilitated by redevelopment of the understory, shrub and sapling layers, which are particularly rapid in broadleaf and mixed forests, providing nesting and foraging habitat for birds33,34. This probably also explains why the greatest divergence in similarity of harvested and reference forests was greater for conifer than for broadleaf or mixed forests.
While we included only studies of clear-cutting (or very low retention), harvesting practices and silvicultural interventions have changed over time; thus, older post-harvest stands might have experienced management that was more, or less, intensive than more recent ones. This could be particularly influential for data representing longer periods post harvest and thus for estimated times to ‘full recovery’. Future post-harvest biodiversity recovery will be strongly subject to climate change effects resulting in the potential for shifts in recovery trajectories4,5,13. Although planting can speed canopy redevelopment, particularly towards conifer dominance, re-establishing mixed woods and structurally complex forests by traditional silvicultural practices has proven challenging35. However, such forests are key reservoirs of biodiversity in boreal landscapes36.
In several cases arthropods, and in some cases lichens and vascular plants, showed a lagged resilience response, such as would be expected due to ecosystem memory5,29 or a lag in local extinction37. For ground beetles and forest floor spiders, such lags could be explained by some mature forest species persisting whereas open-habitat species colonize; with canopy closure at ~15–20 years, open-habitat species disappear while shade species remain at low levels38,39 before eventually increasing to pre-harvest levels by 40–50 years (ref. 38). Lagged resilience for vascular plants in mixed forests could be explained by inclusion of perennial species associated with either a conifer or broadleaf canopy, allowing some species to persist through the changed environment following harvest34,37.
Spiders, birds and in some cases lichens and vascular plants, showed resilience with no lag (that is, substantive post-harvest divergence from the reference forest composition immediately post harvest, followed by a gradual increase). In the case of spiders, this could be explained by Pardosa wolf spiders, which can increase by tenfold within 1–2 years post harvest40. For many bird species, this can be explained by them simply leaving an area when the habitat becomes unsuitable. For vascular plants in conifer and broadleaf forests, this is no doubt due to the direct disturbance of clear-cutting and associated changes in nutrient cycling, light and microclimatic conditions41. Such effects lead to rapid post-harvest increases in shade-intolerant, disturbance-adapted, early successional species accompanied by a rapid decline in shade-tolerant, later-successional species that naturally dominate the understory in conifer stands37,41,42.
Whereas in just over half of cases (biotic groups × forest types) predicted times to ‘full recovery’ of community composition were within 30 years post harvest, important ecological differences remain between harvested and natural forests (for example, characteristics of deadwood, very large trees, population continuity) that could result in longer-term effects on biota21,25,28. Some features of mature forests (for example, large trees and snags, large decayed downed wood) take many decades to re-develop25, delaying recovery of species such as cavity nesters43, saproxylics and epiphytes28,44. As hypothesized, biota that showed no resilience or very long (>95 years) predicted times to ‘full recovery’ (saproxylic beetles, small mammals, bryophytes, lichens) were associated with ecological features that tend to be missing in post-harvest forests (large downed decayed dead wood, large live and dead trees)25,45.
Bryophytes are sensitive to harvesting due to loss of key habitat features, desiccation and increased litterfall associated with broadleaf regeneration46. Their post-harvest recovery can be slow due to their scattered distribution, dispersal limitations and their substrate requirements42,46. For lichens, a long recovery time can be explained by a large set of specialist species inhabiting slowly developing habitat structures, combined with their slow and stochastic recolonization21,44,47. Small mammals had a relatively small decline in community similarity post harvest but no evidence of recovery within 55 years post harvest. This probably reflects species variation in response to harvesting, some being favoured by post-harvest conditions while others decline due to dependence on older-forest features (for example, structural complexity)48. For saproxylic beetles the lack of variation of deadwood during post-harvest succession may be crucial28. Whereas many saproxylic and epiphytic species can persist on logging residues and other residual coarse wood, these habitats tend to decline over time, leading to a lack of large-diameter logs of advanced decay stages, in turn hindering recovery12,25. Our results concur with studies showing that changes in community composition of saproxylic beetles, bryophytes and lichens following clear-cutting lasted more than 50 years (ref. 21).
Our results do not suggest that clear-cut harvest results in forests transitioning to an alternative state, as might be expected due to the loss of important pre-harvest habitat structures49, if tree regeneration failed or composition shifted50 or if a young post-harvest stand burned51. However, such transitions are certainly not precluded in particular cases, indeed we saw many individual examples for which similarity between harvested and reference stands was extremely low, even several decades post harvest (Fig. 4).
There were only three cases of resistance: ‘all arthropods’ in mixed forests, bryophytes and vascular plants in broadleaf forests. The first of these can probably be explained by pooling species with different niches, thus masking variation in responses. For example, a previous study found saproxylic beetles showed lagged resilience while non-saproxylic beetles seemed to demonstrate resistance28. The resistance of bryophytes and vascular plants in broadleaf forests probably reflects the dominance of these understory communities by early successional, disturbance-adapted species34,37,42. Forest floor mosses could survive post harvest due to their high ecological amplitude (for example, survival on remaining tree bases and decaying wood) while decreased seasonal litterfall post harvest enhances bryophyte survival and establishment26,42. Bryophyte species in broadleaf forests are also more adapted to drought and light than those in conifer stands42. Further, broadleaf tree species often regenerate very rapidly post harvest and this can help buffer ground vegetation from the effects of harvest-related soil disturbance and microclimate changes34,42. True epiphytic species will obviously decline sharply with logging as their habitat substrate is removed.
Our results reflect the sampling methods employed in the different studies (Supplementary Table 3). Rare species can easily be missed in standard inventory approaches32,52, and some sampling methods for invertebrates or birds could include individuals using adjacent unharvested forest habitat33. Other than for birds, sampling did not address arboreal biodiversity (for example, epiphytic lichens, bryophytes, arthropods), which can be an important component of biodiversity that presumably increases along with canopy height and diversification21,53,54. Epiphytic lichens could need several hundred years to recover post harvest45. Our reference forest (no recent natural or anthropogenic disturbance) stands were primarily 80–150 years old; only three studies characterized the reference forest as ‘old growth’. We excluded studies where the reference had been heavily managed, but for many European studies reference stands would have been lightly managed or selectively cut long ago21. Thus, we cannot comment on recovery of ‘old growth’ specialists, which comprise an estimated 10–20% of the forest species pool and depend on rarer ecological elements such as large live and dead trees, heterogeneous canopy and large, decayed downed wood21,30,55. Although frequent natural disturbances in boreal forests constrain the development and abundance of such features, a lack of them in managed forests is a major reason for the red-listing of species21,23,28,47,55,56.
The poorer recovery of conifer and mixed, versus broadleaf, forests, and the greater decline in post-harvest similarity between harvested and reference conifer forests, has several possible explanations. Because evergreen conifers with dense foliage and persistent horizontally layered crowns tend to cast heavier shade than deciduous broadleaf trees their removal results in greater microenvironmental changes (that is, light, temperature)57. Furthermore, conifer forests in Europe naturally experience low-severity disturbances resulting in a multi-aged structure, whereas broadleaf forests are more often the product of stand-initiating disturbances19,20. Post-harvest silviculture often aims to return stands to their pre-harvest composition or another desired forest type58. Still, post-harvest canopy redevelopment might be slower in conifer and mixed forests because conifers often require planting and initially grow slowly, whereas broadleaf trees (aspen, birch) have pioneer characteristics such as prolific seed regeneration on disturbed sites, vegetative regeneration and rapid early growth59, perhaps because they grow on more productive sites3. Also, broadleaf forests dominated by aspen or birch are more likely to return directly to a similar composition post harvest20,60, whereas mixed or conifer stands might temporarily have higher broadleaf abundance20,60,61,62.
Post-harvest recovery also reflects the availability and dispersal capacity of species in the regional pool, which, in turn, is affected by natural-disturbance regimes, harvesting footprint and associated fragmentation63,64. Disturbance-driven landscapes, such as the boreal, host biota that are early successional or generalists65; thus, they may have a good supply of colonizers. This could explain why many common boreal songbirds are resilient to moderate harvest levels22. Our results further support this. However, our results cannot provide insight into post-harvest responses for species that function at a larger landscape scale, nor landscape-scale cumulative effects of forest management (fragmentation, age structure, composition) or multiple cycles of harvesting and tending11,12,21,23. Negative impacts of these on biodiversity have been well documented21,66, and forest biodiversity is likely to become increasingly impoverished in landscapes that have a longer history of forest management25,56,64.
Implications for biodiversity conservation
Our results demonstrate clear impacts of harvesting on biodiversity in boreal forests. Although the biotic communities we studied often displayed resilience to clear-cutting, the estimated times to ‘full recovery’ for some biota were longer than the typical time before the next harvest, and there were many individual examples of a complete lack of recovery, even 50–75 years after harvest. Given existing forest rotation cycles, conservation of biodiversity in boreal forests could require extending rotation intervals or managing for ‘old forest’ structural elements retained after harvest (for example, large downed wood, large live and dead trees) and protection of some harvest-age forests from harvesting6. Our focus on similarity of community composition reveals average recovery but does not provide insight for rarer species. Future studies could employ a risk-based or precautionary approach that considers the possibility of reduced abundance or loss of species post harvest (particularly rare, threatened, species of conservation concern, common species) as decreased local diversity can also reduce ecosystem productivity and stability. Conservation of rare species, old-growth specialists and other biota sensitive to stand- or landscape-scale impacts of forest harvesting will require targeted approaches such as natural-disturbance-based management, partial cutting or retention forestry, mixed wood management, continuous cover forestry, multiscale forestry, conservation of biodiversity hotspots and preserving ecosystem memory6,22,67,68.

